Aus der Bukovina im Norden von Rumänien kommt diese Aufnahme einer völlig weißen Nigritellacarpatica. Die Autorin des Buchs The Orchids of Romania, Nora De Angelli, hat die Nigritellacarpatica f. albiflora Ende Juni in der Nähe von Sulița gesehen. Die Blüten dieser seltenen Art im ukrainisch-rumänischen Grenzgebiet sind meist hellrosa bis weißlich – unten rechts eine typische Pflanze. Eine reinweiß blühende Pflanze war bisher nicht dokumentiert.
Photo: Nora De Angelli
Photo: Nora De Angelli
Nigritella carpatica ist diploid und pflanzt sich sexuell fort und ähnelt somit in diesen Eigenschaften Nigritella rhellicani. Zu den Besonderheiten der Art gehören sehr schmale Blätter und kleine Blüten mit einem relativ schmalen Sporn. Die am gleichen Fundort wachsenden Pflanzen zeigen Übergangsformen in der Färbung der Blüten. Möglicherweise gibt es auch bei dieser seltenen Art graduelle Ausprägungen des Pigmentverlusts mit entsprechend allmählichen Übergängen zu der besonderen Albiflora-Form.
Nie zuvor hatte ich eine apochrome Spinnenragwurz gesehen. Das erste Mal ist mir diese besondere Pflanze nun Anfang Mai am nördlichen Gardasee begegnet. Sie hat drei Blüten. Gleich daneben ist noch eine weitere Ophrys spegodes f. albiflora mit einer etwas rundlicheren Lippe, inmitten von etwa zehn Pflanzen mit der vertrauten Färbung einer braun-rötlichen Lippe, Anfang Mai alle leicht abblühend.
Bei Ophrys sphegodes scheint die apochrome Form sehr viel seltener zu sein als etwa bei Ophrys holoserica. Karel Kreutz führt in seinem Beitrag über die „Apochrome Ophrys-Varianten van Midden-Europa“ (1997) vier Standorte in Frankreich und zwei in Deutschland auf.
In der norditalienischen Region Trentino haben die Spinnenragwurze Lippen, die mal rundlicher, mal länglicher sind.
Am nördlichen Ufer des Gardasees liegt der Monte Brione, ein markantes Felmassiv mit Steilhängen nach drei Seiten. Ende April erstrecken sich dort frühlingshaft grüne Wiesen zwischen Olivenhainen und einem von Eschen, Buchen und Steineichen geprägten Wald. In 170 Metern Höhe und damit noch in der unteren Hälfte des 376 Meter hohen Berges, blüht eine weiße Orchis simia, zusammen mit fünf violett blühenden Pflanzen dieser schönen Orchideenart.
Im Regenschauer zeigt sich das weiße Affenknabenkraut in seiner ganzen Schönheit. Die weiß blühende Form ist insgesamt doch sehr selten. Ich schätze d ie Häufigkeit auf zwei Albiflora-Formen je 1000 Pflanzen. Es handelt sich um genetische Besonderheiten, Hinweise auf eine Häufung in bestimmten Regionen oder Biotopen und damit auf eine regional spezifische Entwicklung in der Evolution der Art gibt es nicht.
In einem Beitrag für die Berichte aus den Arbeitskreisen Heimische Orchideen (Peter Zschunke: Von „alba“ bis „viridiflora“ – zur Taxonomie und Biologie hypochromer Orchideen. In: Berichte aus den Arbeitskreisen Heimischer Orchideen 38 (1), 2021. S. 78-99) bin ich der Frage nachgegangen, wie albiflora-Formen von Orchideen wissenschaftlich korrekt bezeichnet werden sollten. In der Vergangenheit waren Botaniker ziemlich kreativ darin, solche Pflanzen zu beschreiben, mit ganz verschiedenen Namen und auf unterschiedlichen Ebenen – bis hinauf zur Subspezies. Dabei erfordert „eine individuelle genetische Beeinträchtigung, wie sie der Ausfall einer Pigmentbildung darstellt, keine Beschreibung eines Taxons“, wie ich in dem Beitrag erkläre. Stattdessen schlage ich vor, „alle Individuen von Orchideen-Arten mit einer ausgeprägten genetischen Störung der Pigment-Bildung auch ohne eine förmliche Beschreibung als albiflora-Formen zu bezeichnen, mit dem meistverwendeten Begriff bei bisherigen Beschreibungen und mit einem Formbegriff in einem allgemeinen Verständnis, der als ’nomen nudum‘ keinen Typusbeleg verlangt“.
Ein Jahr nach der Neuauflage der „Orchideen in Hessen“ von Heinrich Blatt, herausgegeben vom Arbeitskreis Heimische Orchideen (AHO) Hessen, ist jetzt auch erstmals eine Monographie zu den beiden Bundesländern in der näheren und weiteren Nachbarschaft erschienen: „Die wildwachsenden Orchideen in Rheinland-Pfalz und im Saarland„. Von den acht Autoren der Publikation zum 40-jährigen Bestehen des AHO Rheinland-Pfalz/Saarland kamen sechs zur Buchvorstellung nach Koblenz, unter ihnen der AHO-Landesvorsitzende Jürgen Passin (3. von rechts).
Das Rheinland-Pfalz-Buch führt auch etliche albiflora-Formen auf, etwa zu Anacamptis morio, Anacamptis pyramidalis, Dactylorhiza incarnata, Dactylorhiza maculata, Dactylorhiza majalis, Gymnadenia conopsea, Orchis mascula, Orchis militaris und Orchis purpurea. Die Autoren stufen diese als Varietäten ein – botanisch korrekter wäre es, sie als Formen zu bezeichnen. Auch gibt es dabei keine Konsistenz – mal werden sie als var. albiflora, mal als var. alba bezeichnet. Die hypochromen Ophrys-Formen von apifera und fuciflora werden als var. flavescens angesprochen, die von insectifera als var. ochroleuca.
Befremdlich wirkt für mich das Fehlen von Dactylorhiza fuchsii – diese werden unter Dactylorhiza maculata agg. subsumiert. Die Autoren begründen dies damit, dass sich beide Taxa in Rheinland-Pfalz „vermischen“ – während die „saarländischen Populationen … eher dem ‚Fuchsii-Typ'“ entsprechen. Gezeigt werden vor allem Fotos aus der Eifel – diese Pflanzen tendieren eher zu Dactylorhiza maculata mit ihrem nordwesteuropäischen Verbreitungsschwerpunkt. Am Rhein oder im Westerwald wachsen aber auch eindeutige Dactylorhiza fuchsii.
Am Rande der Buchvorstellung und AHO-Mitgliederversammlung gab es viel Gelegenheit zum fachlichen Austausch, etwa mit Karel Kreutz (links) und Werner Hahn (rechts).
Zum Abschluss präsentierten die Buchautoren Jean Claessens und Jacques Kleinen in einem launigen Vortrag einige ihrer detailreichen Supermakro-Fotos und erklärten das präzise Zusammenspiel von Blüte und Bestäubern.
Die Vielfalt der Orchideen-Flora in Rumänien präsentieren Nora De Angelli und Dan Anghelescu in ihrem Buch The Orchids of Romania. Darin finden sich auch 21 Taxa mit Albiflora-Formen – vor allem in den Gattungen Anacamptis, Dactylorhiza und Orchis. Vier weiß blühende Orchideen werden als eigene Varietäten vorgestellt: Dactylorhiza fuchsii var. albiflora, Dactylorhiza incarnata var. albiflora, Dactylorhiza maculata var. albiflora und Epipactis palustris var. albiflora. Der Band ist reich bebildert, mit liebevoll ausgewählten Fotos, darunter etliche mit besuchenden oder bestäubenden Insekten. In Rumänien gebe es 71 Orchideenarten sowie 7 erst kürzlich entdeckte weitere Arten, schreiben die beiden Autoren, die Vater und Tochter sind. Etliche davon seien in umfangreichen Populationen vertreten, auch wenn es zahlreiche Gefährdungen gebe. Wir haben eine moralische Verpflichtung, diese wunderbaren und äußerst verletzlichen Planzen zu schützen, betonen Nora De Angelli und Dan Anghelescu.
Nora De Angelli hat mir auch Fotos von Albiflora-Orchideen in Rumänien geschickt, damit sie hier gezeigt werden können – vielen Dank dafür! Die ersten sind die von Dactylorhiza maculata subsp. transsilvanica, die für das Verstehen von Albflora-Orchideen zwischen spontanen Mutationen und den Wundern der Evolution besonders bedeutsam sind.
Himantoglossum hircinum und Himantoglossum robertianum gehören zu den häufigsten Orchideenarten im südlichen Europa. Immer wieder treten bei beiden Arten Pflanzen auf, deren Blüten das Anthocyan-Pigment fehlt. Diese Albiflora-Formen haben dann grün-weiße Blüten statt der Blüten mit rötlichen Streifen und Punkten.
Foto: Matthias Svojtka, 4.6.2014, in Wien
Matthias Svojtka von der Universität Wien hat auch bei der weitaus selteneren Adria-Riemenzunge, dem Himantoglossum adriaticum, anthocyanfreie Pflanzen beobachtet. „Auf diesem Himantoglossum-Fundort in der Unteren Lobau gibt es eine sehr große Population von ca. 40 – 60 Individuen freistehend auf einer Trockenwiese, stabil zeigen sich auch 1 bis 2 weiße Exemplare„, schreibt er zu diesen seinen Aufnahmen.
Eine zweite Bergwanderung im Corona-Jahr 2020 führt mich Anfang Juli in die Steiermark. Auf der Tauplitz-Alm gilt mein Interesse vor allem der Vielfalt an Nigritella-Arten. Zu den häufigsten Orchideen dieser Bergregion gehört die Mücken-Händelwurz (Gymnadenia conopsea). Sie blühen zu Hunderten auf Wiesen und Waldlichtungen. Auf dem Weg zum Lawinenstein (1965 m) fällt mir am 7. Juli schon von weitem eine weiß blühende Gymnadenia conopsea auf, mit einem lang gestreckten Blütenstand. Diese Art gehört zu den Orchideen, die häufiger weiß blühende Mutationen bilden. Dabei handelt es sich aber meist um einzelne Pflanzen.
In den meisten Teilen ihres Verbreitungsgebiets ist Anacamptis palustris (Synonyme: Orchis palustris, Paludorchis palustris) stark gefährdet, weil es immer weniger Moore und Feuchtwiesen gibt. In einem Niedermoor im bayerischen Landkreis Rosenheim aber wachsen noch Tausende von Pflanzen dieser besonderen Art.
Auf einer Wiese mit etwa 100 blühenden Anacamptis palustris befinden sich auch 7 Albiflora-Pflanzen – das ist ein weit höherer Anteil, als allein aufgrund von genetischen Mutationen zu erwarten wäre. H. Kretzschmar, W. Eccarius und H. Dietrich (Die Orchideengattungen Orchis, Anacamptis, Neotinea. 2007) bemerken: Selten gibt es auch weißblütige Exemplare und bilden das Foto einer Albiflora-Form vom Neusiedlersee in Österreich ab. Es scheint, dass weiß blühende Anacamptis palustris ähnlich selten sind wie bei ihrer Schwesterart Anacamptis laxiflora.
Die Blüten der Albiflora-Pflanzen zeigen nicht den leisesten Hauch eines Purpur-Farbtons. Selbst die Pollinien sind hell, mit einem leicht gelblichen Ton, ähnlich wie bei den Knospen.
Anacamptis palustris wird in diesem Niedermoor begleitet von kleineren Gruppen von Orchis majalis, Dactylorhiza incarnata subsp. ochroleuca, knospenden Epipactis palustris sowie Liparis loeselii.
In einer Population von etwa tausend Orchis pauciflora in Südkroatien hat Bariša Ilić aus Metković acht Pflanzen mit fast weißen Blüten gefunden. Die roten Punkte auf der Lippe bilden so einen besonderen Kontrast. Es war das erste Mal, dass er diese besonderen Formen an dem Fundort in der Nähe der Stadt Ston im Bezirk Dubrovnik-Neretva gesehen hat. Bariša Ilić ist ein Ornithologe, der sich – wie er schreibt – vor fünf Jahren in wild wachsende Orchideen verliebt hat.
Orchis pauciflora – photo: Bariša Ilić
Die Blüten von Orchis pauciflora haben eine kräftige gelbe Farbe. Sie enthalten hohe Anteile von Carotinoiden – den Pigmenten, die bei vielen Pflanzen die gelbe Blütenfarbe erzeugen. Während Anthocyanine – die rote, purpurne oder bräunliche Blütenfarben ermöglichen – wasserlöslich sind, sind Carotinoide fettlöslich (wie Chlorophyll). Da Anthocyanine in einem komplexen Prozess der Biosynthese gebildet werden, an dem mehr als fünf Enzyme beteiligt sind, ist es bei Anthocyaninen wahrscheinlicher, dass es zu einem Pigmentverlust kommt, als bei Carotinoiden. Dies könnte erklären, warum Orchis pauciflora mit weißlichen anstelle von gelben Blüten äußerst selten sind. Und selbst bei diesem Fund scheinen die Pflanzen im Lippenzentrum noch einen Rest von Gelb erhalten zu haben.
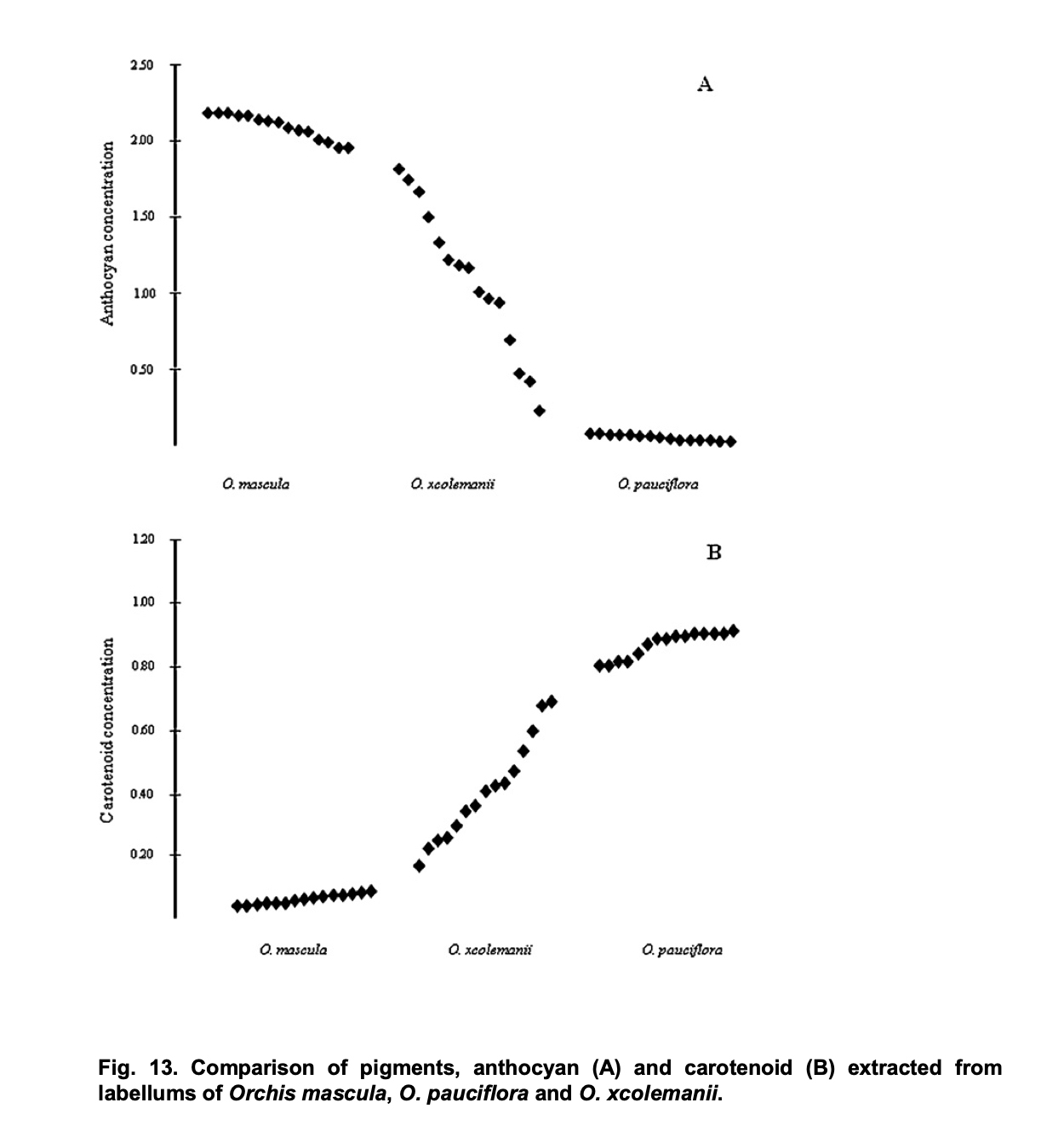
Dieser Eindruck eines graduellen Verlusts von Carotinoiden passt zu einer interessanten Studie der italienischen Botanikerin Alessia Luca. Diese untersuchte die Konzentration von Carotinoiden bei Hybriden von Orchis pauciflora und Orchis mascula. Ihre Doktorarbeit an der Universität von Kalabrien enthält Messungen der Anthocyanin- und Carotinoid-Konzentrationen in der Lippe der als Orchis x colemanii bezeichneten Hybriden. Dabei zeigte sich, dass die Carotinoid-Konzentration bei den Hybriden allmählich abnimmt: „O. xcolemanii showed a continuous flower color variation (Figure. 13) ranging from red-purple flowers of O. mascula to yellow flowers of O. pauciflora.“ (Alessia Luca: Evolutive significance of hybridization in Mediterranean deceptive orchids, p. 42)
Alessia Luca: Evolutive significance of hybridization in Mediterranean deceptive orchids, p. 43