Warum sind Ophrys-Blüten mit einer Pigmentstörung gelblich-grün und nicht weiß wie bei Orchis, Dactylorhiza oder anderen Gattungen? Der Antwort auf diese Frage bin ich auf der Orchideentagung in Sundheim nachgegangen, in einem Vortrag mit dem Titel: Die Ophrys-Blüte – mehr als eine Einladung zur Pseudokopulation.

Meine These: Die Lippen aller Ophrys-Blüten sind grün – wir können das aber meist nicht sehen, weil das Blattgrün in den Lippen vom Anthocyanin überdeckt wird. Erst wenn die Anthocyanin-Produktion gestört ist, wird in der Ophrys-Lippe das Chlorophyll sichtbar, zusammen mit ebenfalls meist enthaltenen gelben Pigmenten. Nur die Lippenzeichnung ist frei von Pigmenten, sie erscheint bei einem Ausfall der Anthocyanin-Bildung meist weiß. Aber selbst in den unteren Schichten des bläulich schimmernden Mals von Ophrys speculum haben Forscher nachgewiesen, dass sich dort Chloroplasten befinden, also Zellpartikel mit Chlorophyll. Die bläuliche Farbe resultiert aus den Vakuolen der Epidermis, die mit Cyanidin-Pigmenten gefüllt sind, während der bräunliche übrige Teil der Lippe zusätzlich auch Delphinidin und Quercetin-Pigmente sowie Anthocyanine enthält.
Bei den Ophrys-Indidviduen mit einem Ausfall der Pigment-Bildung wäre die Bezeichnung weiß blühend oder albiflora fehl am Platz. Besser passt der Begriff hypochrom in der Bedeutung einer unterentwickelten Farbpigmentausprägung.
Belege dafür gibt es bei mehr als 20 Ophrys-Arten.
Eine ganze Population von Ragwurz-Pflanzen ohne Anthocyanin in der Lippe gibt es bei Basel von Ophrys apifera. Diese Pflanzen sind auch als Basiliensis-Varietät beschrieben.
Anthocyanin dient auch als Sonnenschutz, es hat die Eigenschaft, schädliche Lichtenergie wie etwa die UV-Strahlung zu absorbieren. Die wasserlöslichen Anthocyanin-Pigmente befinden sich in den Vakuolen der Pflanzenzelle, die auch Wasser und Nährstoffe enthalten. Besonders die Pflanzen in Südeuropa sind auf die Sonnenschutz-Funktion des Pigments angewiesen.
Im allgemeinen haben die Petalen höherer Blütenpflanzen kein Chlorophyll. Das Gewebe ihrer Blüten und Früchte enthält keine Chloroplasten – also Zellpartikel, die Chlorophyll enthalten und deswegen grün gefärbt sind – sondern Chromoplasten. Diese enthalten kein Chlorophyll, sondern zum Beispiel Carotenoide: gelbe, orangene oder rote Farbpigmente.
Nur im Knospenstadium haben einige Blütenpflanzen noch höhere Anteile von Chloroplasten – diese entwickeln sich dann zu Chromoplasten oder farblosen Leukoplasten mit der Funktion, Eiweiß oder Stärke zu speichern – hierzu gibt es einen interessanten Beitrag von Kevin Pyke und Anton Page. Die gleiche Umwandlung lässt sich beim Reifen von Tomaten beobachten – die zunächst grüne Frucht hat noch viele Chloroplasten mit Chlorophyll, die sich dann zu Chromoplasten mit dem roten Pigment Lycopin umwandeln.
In Laubblättern enthält eine Blattzelle etwa 20 bis 50 durch Chlorophyll grün gefärbte Chloroplaste. Sie sind mit Stroma gefüllt, das auch das Enzym Ribulose-Bisphosphat-Carboxylase/Oxygenase (RuBisCo) enthält. Dieses leistet im Zusammenwirken mit Wasser und Sonnenlicht einen entscheidenden Beitrag für die Umwandlung von Kohlendioxid in Glucose: Aus CO2 und H2O wird C6H12O6.
Forschungen haben gezeigt, dass auch Orchideen sonniger Wiesenstandorte im Mittelmeerraum zumindest als partiell mykoheterotroph zu betrachten sind, also einen Teil ihrer Nährstoffversorgung von Pilzen erhalten. Weil Pilze den für die Enzymbildung elementaren Stickstoff im Boden sehr viel besser lösen können als höher entwickelte Pflanzen, sind mykoheterotrophe Orchideen besser mit Stickstoff versorgt als Pflanzen anderer Familien ohne Nährstoffversorgung durch Pilze. Die einzelnen Orchideenarten verbinden sich mit jeweils unterschiedlichen Pilzen, bei vielen Orchideen lassen sich Bodenpilze der Gattung Tulasnella als Versorger feststellen.
Zusätzlich aber nutzen Ophrys-Orchideen auch den Umstand, dass Chloroplasten im Blütengewebe erhalten sind, so dass hier weiter Photosynthese stattfinden kann. Einige Ophrys-Arten haben auch ein intensiv grünes Perigon – dort sind dann anders als in der Lippe keine Anthocyanine enthalten, die das Chlorophyll überdecken würden.
Dabei nutzen Ophrys-Blüten, so ist zu vermuten, den Crassulaceen-Säurestoffwechsel, nach der englischen Bezeichnung Crassulacean Acid Metabolism als CAM abgekürzt. Das ist ein nach der Familie der Dickblattgewächse, der Crassulaceae, benannter Mechanismus der Photosynthese, der besonders an heiße und trockene Standorte angepasst ist.
Der Vorteil dieser CAM-Photosynthese gegenüber der gängigen C3-Photosynthese: Die Pflanzenzellen nehmen Kohlendioxid nachts auf. Die Spaltöffnungen im Pflanzengewebe sind dann geöffnet. Und in der Tageshitze können sie geschlossen bleiben, so dass die Pflanze vor Austrocknung geschützt ist. Das nachts aufgenommene CO2 wird in den Vakuolen der Zelle in Form von Äpfelsäure gespeichert und tagsüber mit Lichtenergie in Kohlendioxid und Sauerstoff umgewandelt. Die CAM-Photosynthese begnügt sich im Vergleich zur C3-Photosynthese mit weniger als einem Fünftel der benötigten Wassermenge. Somit kann die Pflanze in Trockenzeiten gut überleben, bleibt vor Stress wegen Wassermangels geschützt.
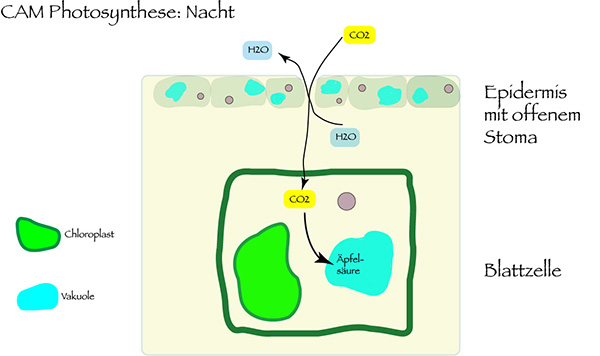
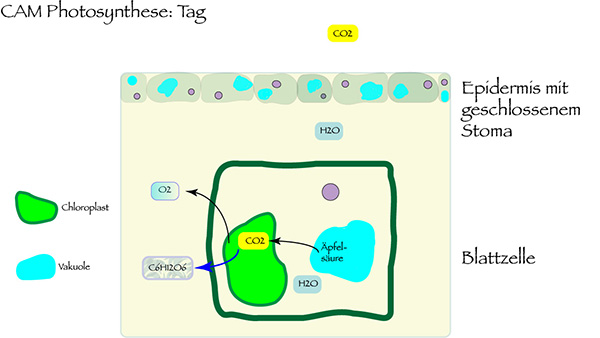
Es gibt auch Pflanzen mit fakultativer CAM-Photosynthese – hier wird der Wassersparmodus nur bei sommerlicher Trockenheit eingeschaltet. Im Frühling setzen die knospenden Pflanzen dann noch die C3-Photosynthese ein. Denn CAM hat neben dem Vorteil des geringeren Wasserverbrauchs einen großen Nachteil: Die Produktion von Biomasse fällt deutlich geringer aus. CAM-Pflanzen wachsen sehr viel langsamer als C3-Pflanzen.
Orchideen mit dünnen Laubblättern haben in der Regel eine C3-Photosynthese – dazu gehören etwa die Gattungen Orchis, Dactylorhiza, Anacamptis und Neotinea. Solche mit dicken Blättern wie die tropischen Gattungen Phalaenopsis, Cymbidium oder Cattleya praktizieren die CAM-Photosynthese, also den Crassulaceen-Säurestoffwechsel.
Da die CAM-Pflanzen das nachts aufgenommene CO2 für die tagsüber ablaufende Photosynthese speichern müssen, haben sie oft stark vergrößerte Vakuolen. Dicke Blätter können mehr organische Säure speichern, die das CO2 über Nacht konserviert. Es könnte sein, dass dies auch bei der Blütenblatt-Photosynthese von Ophrys der Fall ist. Ein Indiz dafür ist die verdickte, fleischige Ophrys-Lippe – und diese Form kommt wiederum der Einladung an die Bestäuber zur Pseudokopulation entgegen. Auch die oft geringe Wuchshöhe ist ein Indiz dafür, dass Ophrys-Arten zumindest fakultative CAM-Pflanzen sind – es wird weniger Biomasse aufgebaut.
Ein Vorteil der floralen Photosynthese ist die ideale Position der Blüten zum Sonnenlicht hin. Die höhere Aufnahme von Lichtenergie ermöglicht eine höhere Kapazität zum Speichern von CO2. Auch wenn es noch keine eingehenden Forschungen dazu gibt, sprechen viele Hinweise dafür, dass insbesondere die mediterranen Ophrys-Arten eine florale Photosynthese beherrschen. Mit dem zumindest fakultativen CAM-Stoffwechsel verfügen sie über die Fähigkeit, auch in trockenem Klima, auf Felsgrund und mit früh welkenden Laubblättern zu bestehen. Die Entwicklung der Ophrys-Lippe ist damit gleich zwei funktionalen Zielen gefolgt, der Anpassung an einen Bestäuber und der Befähigung zu floraler Photosynthese.
Damit zeichnet sich im Vegetationszyklus der Ophrys-Pflanze eine idealtypische Abfolge von Phasen ab, die jeweils an spezifische Umweltbedingungen und biologische Anforderungen angepasst sind. Für einzelne Arten und Standorte gibt es sicherlich Unterschiede, generalisierend lassen sich aber diese Phasen unterscheiden:

Der Keimling entwickelt sich aus der mykoheterotrophen Nahrungsversorgung und erhält mit der Ausbildung seiner ersten Blattrosette die Befähigung zur Photosynthese. Es ist anzunehmen, dass hier der C3-Mechanismus der Photosynthese vorherrscht, im Frühling sind die Temperaturen noch gemäßigt und es steht genug Feuchtigkeit zur Verfügung. Auch geht es hier für die Pflanze vor allem um schnelles Wachstum, den zügigen Aufbau von Biomasse. Dabei ist die C3-Photosynthese klar im Vorteil.
Die knospende Pflanze ist weiter auf Wachstum angelegt, die Blätter erreichen ihre optimale Kapazität zur C3-Photosynthese.
Mit der Blüte beginnen die Laubblätter zu welken, ergänzend übernimmt die Blüte die CAM-Photosynthese, und sichert so trotz zunehmener Hitze und Trockenheit eine ausreichende Nahrungsversorgung.
In der Fruchtreife ist mit dem Anschwellen der Fruchtknoten noch einmal der Aufbau von Biomasse erforderlich. Mit dem Abblühen verringert sich die Kapazität der Photosynthese. Möglicherweise gewinnt jetzt auch die mykoheterotrophe Nahrungsversorgung wieder an Bedeutung.