Saure Moore sind nicht gerade ein typischer Standort für Orchideen – aber es gibt zwei seltene Ausnahmen: Die eine erstreckt sich über ein geographisch ausgedehntes Gebiet von Belgien über Nordwestdeutschland bis Skandinavien. und wird meist als Dactylorhiza sphagnicola bezeichnet. Die andere wächst nur in der dänischen Region Thy: Wenige hundert Meter hinter der Küstenlinie der Nordsee gibt es eine Population weiß blühender Orchideen, die von Henrik Ærenlund Pedersen als Dactylorhiza majalis subsp. calcifugiens beschrieben wurden (in: Nordic Journal of Botany, 2004). 2007 haben dann Sebastian Sczepanski und Karel Kreutz erklärt, dass es angemessener wäre, diese Pflanzen als Subspezies von Dactylorhiza sphagnicola zu betrachten – während Pedersen und Mikael Hedrén sphagnicola lediglich als weitere Subspezies von Dactylorhiza majalis betrachten. Abgesehen von der Farbe sind die morphologischen Unterschiede der Einzelblüten von Dactylorhiza majalis subsp. majalis (link), calcifugiens (Mitte) und sphagnicola (rechs) kaum erkennbar:
 Der Sporn von Dactylorhiza sphagnicola ist ein wenig länger als der von D. majalis subsp. calcifugiens. Und die Blätter dieser Art sind in einem größeren Winkel zur Seite gespreizt als bei D. sphagnicola:
Der Sporn von Dactylorhiza sphagnicola ist ein wenig länger als der von D. majalis subsp. calcifugiens. Und die Blätter dieser Art sind in einem größeren Winkel zur Seite gespreizt als bei D. sphagnicola:

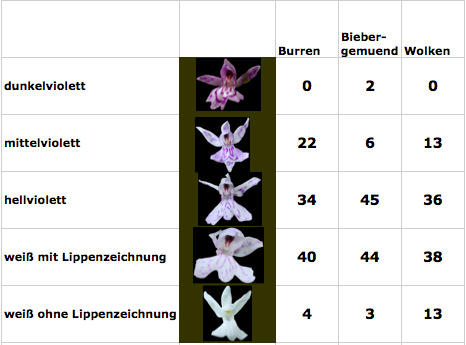
Die Blüten zeigen keinerlei rosa oder violetten Farbton, selbst den Pollinarien fehlt jedes Anthocyanin. Im Zentrum der Blüte, an der Narbenhöhle, findet sich eher ein leicht gelbliche Färbung, was ein wenig an Dactylorhiza incarnata subsp. ochroleuca erinnert. Im Unterschied zu anderen Populationen von Albiflora-Formen, etwa bei Dactylorhiza fuchsii, gibt es hier auch keine graduellen Unterschiede beim Verlust der Farbe – alle Pflanzen sind konsistent in der weißen Farbe ihrer Blüten.

Bei der Erkundung der Region, habe ich Calcifugiens-Pflanzen an zwei Standorten gefunden, einmal in der Nähe des kleinen Fischerdorfs Lild Strand mit nur drei Pflanzen, dann weiter südlich in einem Moor namens Nissekaer mit etwa 150 Pflanzen. Umgeben von Dünen ist dieser Ort eine natürliche Senke mit einer Länge von etwa 1500 und einer Breite von etwa 250 Metern:

Mitte Juni sind die Orchideen hier gerade am Anfang ihrer Blütezeit. Die meisten Dactylorhiza majalis subsp. calcifugiens wachsen am Rand der feuchten Stellen, nicht so nass stehend wie Dactylorhiza sphagnicola im Hohen Venn in Belgien. Und die Calcifugiens-Pflanzen sind kleiner, erreichen gerade mal eine Höhe von 31 cm. Zu den Nachbarpflanzen gehören Torfmoos (Sphagnum palustre), Teich-Schachtelhalm (Equisetum fluviatile), Wollgras (Eriophorum angustifolium), Fieberklee (Menyanthes trifoliata), Moosbeere (Vaccinium oxycoccus), Besenheide (Calluna vulgaris), Siebenstern (Trientalis europaea) und sogar Sonnentau (Drosera rotundifolia). Unter den niedrig wachsenden Büschen dominiert der Gagelstrauch (Myrica gale), der von der Bierbrauerei in der nahegelegenen Stadt Thisted zum Bierbrauen verwendet wird.

Einige Calcifugiens-Pflanzen haben eine breitere Lippe, was einen möglichen Hybrid-Einfluss von Dactylorhiza maculata andeutet – ähnlich wie bei den Sphagnicola-Pflanzen im Hohen Venn.

Unter all den weiß blühenden Orchideen im Nissekaer-Moor fand ich zwei violett blühende Pflanzen: Die eine möglicherweise eine Hybride von Dactylorhiza majalis subsp. calcifugiens und Dactylorhiza maculata (links), die andere vermutlich eine noch knospende Dactylorhiza maculata (rechts):

Als Besucher und möglicherweise Bestäuber von Dactylorhiza majalis subsp. calcifugiens habe ich eine Schwebfliege der Gattung Syrphida beobachtet – wie ich es zuvor auch bei Dactylorhiza sphagnicola im Hohen Venn (links) gesehen habe.