The Nepalese orchid expert Bhakta Bahadur Raskoti found an albiflora form of Neottianthe cucullata together with its regular form. This orchid is distributed in central and western Nepal at altitudes of 3700 to 5000 m. It’s flowering in August. The albiflora form has been described in 1995 by the Chinese botanist P.Y. Fu. The species has its western distribution limit in Poland.
Before the end of the year I received special finding reports from Lithuania: Bernd Gliwa has found a white-flowered Dactylorhiza traunsteineri, only once in 2006. The location is in a lime marsh between Kaunas and Šiauliai. It’s quite remarkable that this Dactylorhiza species is quite rare to see with white-flowered forms.
Photo: Bernd Gliwa, 21/06/2011, near Dievogala/Lithuania
In this nature reserve he has also photographed this Dactylorhiza incarnata. In this area there are also Dactylorhiza incarnata subsp. ochroleuca (with W. Eccarius, Die Orchideengattung Dactylorhiza, 2016 it’s again a species of its own, D. ochroleuca), Dactylorhiza incarnata subsp. cruenta and Dactylorhiza majalis subsp. baltica.
Bernd Gliwa has also studied the dragonflies of Lithuania and just published a monograph about them: Lietuvos laumžirgiai.
While travelling in Attica, Greece, Marco Klüber photographed this special Ophrys helenae which demonstrates the bilateral symmetry of orchid flowers. The left half of the lip shows the regular red-brown coloring while the right half is hypochrome, with a partial loss of pigments. The lighter red hues are preserved, in addition to the chlorophyll in the lower edges.
Orchid flowers have a bilateral symmetry – as it’s the case with beetles or the human face elsewhere in nature. Other plants like the flowers of liliaceae have a radial symmetry with three or more mirror lines.
On his trip Marco also saw the albiflora form of Anacamptis papilionacea subsp. messenica (formerly subsp. heroica):
Following the benchmark books about the genera Anacamptis, Neotinea and Orchis (together with Horst Kretzschmar und Helga Dietrich, 2007) as well as a monograph about the genus Cypripedium (2009), Wolfgang Eccarius now has finished his long lasting work and has published a compendium about the Dactylorhiza orchids.
The book closes a big gap and covers a genus of orchids which is especially rich in species and quite widespread in Europe. As he has established in former publications, the author at first gives a comprehensive introduction before portraying 37 species and 46 subspecies. Eccarius explains his methodological approach and offers a summary of the research history, beginning with the plant book of Otto Brunfels published in 1534. The book abstains from giving a system to identify the species by morphological indicators. But a tree of the genus structure based on genetical research offers a good overview about the manifold Dactylorhiza orchids and their relations.
In his description, Eccarius has taken some main decisions. He abstains from describing varieties and forms, arguing that those terms are “highly problematic” with Dactylorhiza. “The main goal of the author was a genus structure wich matches logical principles as well as the needs of observations in the field.” He stresses that’s it’s more important to differentiate between the ten sections than defining species: “With Dactylorhiza, sections are much easier to define than species.” For example, the Fuchsiae form a section of their own, together with Dactylorhiza saccifera. The section of Majales comprise Dactylorhiza majalis, Dactylorhiza cordigera and Dactylorhiza elata.
Photo: Peter Zschunke, 16/06/2012, bei Glæde, Dänemark
It’s comprehensible that Eccarius views Coeloglossum viride as Dactylorhiza viridis. The Viridae are presented as a subgenus, in addition to the subgenus Dactylorhiza. Other taxonomic decisions are more thought provoking. For example when the yellowish Early Marsh Orchid no longer is a subspecies of Dactylorhiza incarnata, but an own species Dactylorhiza ochroleuca – because there are only very few hybrids between both “which justifies the treatment as a single species in the view of the author”.
Difficult are the explanations about the white flowering Dactylorhiza fuchsii in Ireland, which are elevated by Eccarius to the status of a subspecies – while most experts view the okelly taxon of Dactylorhiza fuchsii as a variety. And the morphological description of the author is not quite helpful in the field: “The subspecies is different by its lower growth” – while the photos show rather high plants. And: “The white color of flowers is shown by whole populations and not only by single plants.”
But this is also valid for Dactylorhiza majalis subsp. calcifugiens, which is presented by Eccarius only as a synonym to Dactylorhiza sphagnicola. The book shows a photo of a plant from the German region of Celle which seems to be an albiflora form of Dactylorhiza sphagnicola, but which is quite different form the calcifugiens population in Northern Denmark.
Quite useful are the explanations about Dactylorhiza maculata, which is presented as a west and northern European species, distributed also in Northern Africa and Northern Asia. The color of flower is described as especially variable, from pure white to a soft and light purple.
Eccarius understands the tendency to a color dimorphism (or polymorphism) which is typical for the genus as functionally relevant. With Dactylorhiza romana, sambucina or incarnata this phenomenon serves as a factor, “to avoid quick learning experiences of polinating insects”. This matches with the regionally different tendency of Dactylorhiza fuchsii to develop albiflora forms.
The new book makes big progress in understanding the Dactylorhiza orchids. But for a full perception there is still a lot of research necessary.
Why are Ophrys flowers with a defect in pigment production yellowish-greenish and not white as it’s the case with Orchis, Dactylorhiza or other genera? I’ve tried to give an answer to this question at the orchids conference in Sundheim (Germany), in a lecture titled The Ophrys flower – more than an invitation to pseudo copulation.
Ophrys holoserica
My assumption: The lips of all Ophrys flowers are green – but in most cases we can’t see it, because the chlorophyll in the lips is covered by anthocyanins. Only if the production of anthocyanins is disturbed, the chlorophyll in the Ophrys lip is visible, often together with yellow pigments. Only the marking of the labellum is free of those pigments, it appears in the case of a defect in Anthocyanin production as white. But even in the lower segments of the bluishly shining marking of Ophrys speculum researchers have showed the existence of chloroplasts, of cell plastids with chlorophyll. The bluish color comes from the vacuoles of the epidermis which are filled with cyanidin pigments, while the brown rest of the labellum in addition contains delphinidin, quercetin and anthocyanin pigments.
The Ophrys forms without anthocyanins cannot be termed as white-flowered or albiflora. It makes more sense to call them hypochromic in the meaning of a underdeveloped production of color pigments.
Those forms have been found with more than 20 Ophrys species.
Near the Swiss town of Basel there is a whole population of Ophrys apifera plants without anthocyanins in the labellum. Those plants have been described as Ophrys apifera var. basiliensis.
Anthocyanins also acts as a sun protection. They have the ability to absorb damaging light energy as UV UV radiation. The water-soluble anthocyanin pigments are in the vacuoles of the plant cell, which also contain water and nutrients. This protective function of the pigment is especially important for plants in Southern Europe.
In general, the petals of flowering plants don’t have chlorophyll. The tissue of their flowers and fruits don’t have chloroplasts (those cell parts which contain chlorophyll and therefore are green), but chromoplasts. Those don’t have chlorophyll, but for example carotenoids: yellow, orange or red color pigments.
Only in the early bud stadium some flower plants still have some amounts of chloroplasts – later, those develop to chromoplasts or colorless leucoplasts with the function to store proteins or starch – this process has been explored in an interesting report by Kevin Pyke and Anton Page. The same transformation can be observed when tomatoes are ripening – the fruit is green at first, with many chloroplasts and chlorophyll, which later change to chromoplasts with the red lycopene pigment.
The cells of green leaves have about 20 to 50 chloroplasts with chlorophyll. They are filled with stroma, which also contains the enzyme Ribulose-Bisphosphat-Carboxylase/Oxygenase (RuBisCo). Together with water and sun light, this enzyme has a decisive role in transforming carbon dioxide in glucose: from CO2 and H2O to C6H12O6.
Studies have demonstrated that orchids of sunny meadows in the Mediterranean can be viewed as mycoheterotrophic: They get at least a part of their nutrients from fungi. Since fungi can dissolve the nitrogen (which is important to produce enzymes) of the soil much better than higher developed plants, mycoheterotrophic orchids have a better nitrogen supply than other plants without this special relationship. The different orchid species have different relationships with fungi, but many orchids rely on fungi of the genus Tulasnella.
Additionally, Ophrys orchids make use of the fact, that they have chloroplasts in their flower tissue, so photosynthesis becomes possible in the flower. Some Ophrys species also have intense green petals – in contrast to the labellum there are no anthocyanins in these petals which would otherwise cover the chlorophyll.
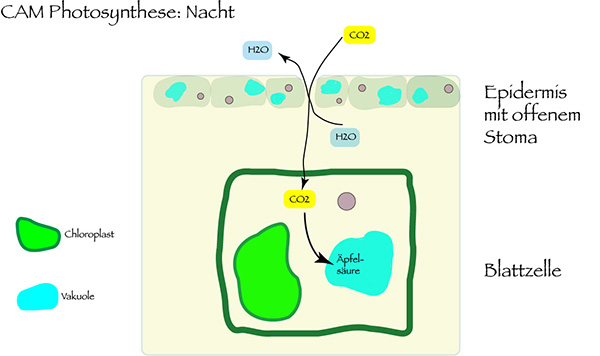
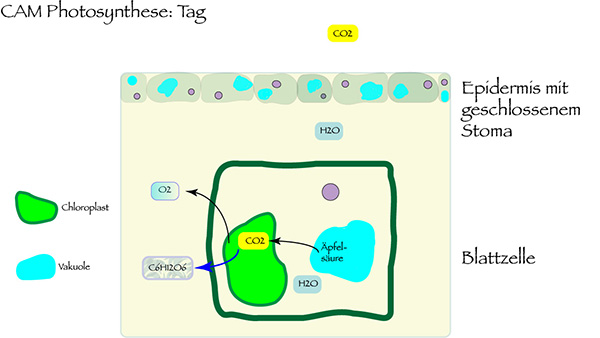
It can be assumed that Ophrys flowers make use of the Crassulacean Acid Metabolism (CAM). This mechanism of photosynthesis, named after the family of the Crassulaceae, is especially adapted to hot and dry places.
The advantage of this CAM photosynthesis compared with the standard C3 photosynthesis: The plant cells receive carbon dioxide at night. Then, the stomata of the plant tissue are open. In the daily heat they can be closed, so the plant is protected from dehydration. The CO2 received by night is stored in the vacuoles of the cell in form of malic acid. By day, it will be transformed to carbon dioxide and oxygen by the help of light energy. The CAM photosynthesis only needs less than a fifth of the water amount which is normally necessary in the case of C3 photosynthesis. Therefore, the plant can survive in dry times and is protected against lack of water.
There are also plants with a facultative CAM photosynthesis: the water saving mode will only be used in dry summer times. In spring, the budding plants still use the C3 photosynthesis. While CAM has the advantage of a lesser water consumption it also has a big disadvantage: The production of biomass is quite lower. CAM plants grow much slower than C3 plants.
Most orchids with thin leaves use a C3 photosynthesis. Among them are the genera Orchis, Dactylorhiza, Anacamptis and Neotinea. Those with thick leaves can master CAM photosynthesis, for example the tropical genera Phalaenopsis, Cymbidium or Cattleya.
Since CAM plants have to store CO2 by night in order to enable the daily photosynthesis, they have enlarged vacuoles. Thick leaves can store more organic acid which conserves CO2 in night time. It may well be that this is also the case with the floral photosynthesis of Ophrys. One clear sign is the thick Ophrys labellum – the form which also has its special function to imitate female pollinators to induce pseudo copulation. The often low height of Ophrys is an additional sign that those species are at least facultative CAM plants, since there is less biomass produced.
An advantage of the floral photosynthesis is the ideal position of flowers to the sun light. The higher amount of light energy absorbed enables a higher capacity to store CO2. Though there is no detailed research yet, there are many signs that especially the Mediterranean Ophrys master a floral photosynthesis. The CAM metabolism enables them to survive in dry climates, on rock grounds, with early withering leaves. The evolution of the Ophrys labellum fulfilled two functions: the adaption to pollinators and the ability of floral photosynthesis.
A cycle of vegetation becomes visible which determines the life of Ophrys. There are four phases which are adapted to specific environment conditions and biological demands:
The seedling is developing under conditions of a mycoheterotrophic supply with nutrients. Developing the first leaves, photosynthesis becomes possible. It can be assumed that the C3 mechanism of photosynthesis is still dominant, since temperatures are moderate in spring and there is enough water. And the plant needs to grow quickly, to build up biomass. Here the C3 photosynthesis has a clear advantage.
The budding plant is still growing, the leaves reach their optimal capacity for C3 photosynthesis.
Beginning with flowering the leaves begin to wither. So the flower starts its additional CAM photosynthesis and secures a sufficient supply with nutrients even under conditions of increasing heat and drought.
When the fruits ripe there is again a phase when biomass has to be added. But the capacity of photosynthesis is decreasing when the flower is withering. Now the role of the mycoheterotrophic supply with nutrients might become more important again.
The 20th conference of orchid experts in Kehl-Sundheim invited its participants to a long journey: The lectures had a broad range from the German regions of Odenwald and the upper valley of the Fils to Austria and Iran.
Orchids Conference Sundheim
The trip report form Iran opened the conference: Jean-Marc Haas showed impressive pictures of orchids, tulips and Fritillaria. Markus Sonnberger from Heiligkreuzsteinach presented a botanical profile of the Odenwald. Among orchids, Himantoglossum hircinum and Epipactis helleborine are increasingly present, while other species are declining: Dactylorhiza fuchsii, Dactylorhiza majalis and Orchis mascula. In this region there are also growing Cephalanthera longifolia, Orchis militaris, Neottia ovata, Neotinea ustulata and Platanthera bifolia. Markus Sonnberger also showed botanical rarities of the region as Buxbaumia viridis or Stellaria neglecta.
H. Moeller showed impressive pictures of potential pollinators of Neottia ovata, among them wasps of different sizes. He observed them at short distance with his Lumix compact camera.
Norbert Griebl lectured about the “finest orchid regions of Austria” some albiflora forms as Gymnadenia conopsea in white and greenish, at the Golzentipp in the Gailtal Alps, or a white flowering Anacamptis coriophora in the valley of Lobau.
Prof. Hannes Paulus from Vienna presented the latest results of his research about the pseudo copulation of Ophrys species. He criticized a “unprecise use of the subspecies term in botany, that’s a big mess”. From his view it’s not correct to describe Ophrys illyrica und Ophrys tommasinii as subspecies of Ophrys sphegodes – “both have a different size and different pollinators, they are different species”.
Helmut Zelesny viewed white flowering forms of Orchis militaris, Gymnadenia conopsea and Neottia nidus-avis in the upper valley of the Fils near Unterboehringen as “freaks of nature, without scientific value”. But this does not explain why white flowering forms of some species and in some regions are more common than in other cases. Zelesny also showed the photo of a hybrid of a white flowering Orchis mascula and Orchis pallens.
Bees and other insects are looking for floral rewards, such as nectar and pollen. And they learn to associate a variety of floral cues, including color with such rewards, as three researchers of the University of Arizona – Avery L. Russell, China Rae Newman and Daniel R. Papaj – explain in an article published this year in Evolutionary Ecology (DOI 10.1007/s10682-016-9848-1). Yet, we don’t know much about the ways how insects are learning.
Andrea spec. with Orchis simia
The Arizona project looks for plants with different colors due to a single loss-of-function mutation blocking the production of floral pigments. Similar to the albiflora orchids presented on this web site, such color polymorphisms may also occur in other plant families. The authors explain that those are quite common, for example with Geranium thunbergii, Antennaria dioica or Aquilegia coerulea – a relative to the European Aquilegia vulgaris.
Aquilegia vulgaris
There study analyzes the behavior of pollinators of hypochromic Solanum tridynamum in an experimental arrangement. It could be shown that initially naïve bees had no preference for purple- or white-flowered plants. The same was the case when the researchers prevented the release of pollen – this was done by sealing the anther pore with glue. But bees with a rewarding experience on plants with purple corollas expressed strong, significant landing preferences for morphs with purple corollas relative to morphs with white corollas. This preference to the rewarding color was much weaker in the case of rewarding flowers with white corollas. The authors came to the conclusion: bees showed a bias in terms of how experience shaped preference: experience with the purple morph had a greater effect on preference than experience with the white morph.
The reason for this bias in learning might be, according to the Arizona researchers, that purple flowers exhibit much greater chromatic contrast, so the hypochromic Solanum tridynamum is likely even less discriminable in real-world foraging conditions. Such biases, the authors conclude, might curtail the success of such morphs and perhaps even contribute to the low frequencies in which they occur. The Arizona case study titled White flowers finish last: pollen-foraging bumble bees show biased learning in a floral color polymorphism is great research – there should be others to follow to see if pigmented flowers have an a priori advantage over hypochromic flowers.
Photo: Christian Schlomann, 28/05/206, Campo Imperatore/Abruzzo, Italy
Christian Schlomann has visited the Abruzzo mountains and sent me this beautiful Orchis purpurea. Campo Imperatore is a mountain grassland in the Gran Sasso massiv, in an altitude between 1500 and 1900 meters.
Ophrys flowers which lack Anthocyanin pigments often have a green labellum, with a white pattern. This is beautifully demonstrated by this Ophrys bertolonii in Southern Croatia. It is quite similar to the one which Pavel found in Northern Croatia.
The first albiflora contribution of this spring is coming from Werner Hahn who has found and photographed this impressive Anacamptis collina in the Southern Spanish region Extremadura.