This photo of a totally white-flowered Nigritellacarpatica was taken in the region of Bucovina in Northern Romania. End of June, the author of the book The Orchids of Romania, Nora De Angelli, has seen this Nigritellacarpatica f. albiflora near of Sulița. The flowers of this rare species in the region between Ukraine and Romania are mostly bright pink to whitish. Below there is a typical plant on the right. Up to now, a totally white-flowered plant was not known.
Photo: Nora De Angelli
Photo: Nora De Angelli
Nigritella carpatica is diploid and has a sexual reproduction – in these characteristics the species is similar to Nigritella rhellicani. The species is characterized by very narrow leafs and small flowers with a relatively narrow spur. The flowers of other plants at this location show gradually changing color hues. This rare species probably has a gradual loss of pigments with smooth transitions to the special albiflora form.
Never before I had seen an apochrome spider-orchid. Now I have encountered this special plant for the first time in the beginning of May at the Northern edge of lake Garda. She has three flowers. At her side is another Ophrys spegodes f. albiflora with a more rounded lip, surrounded by about ten plants with the common hues of a brown-reddish lip, most of them already withering.
The apochrome form of Ophrys sphegodes seems to be more rare than with Ophrys holoserica. Karel Kreutz mentions four places in France and two in Germany in his paper about Apochrome Ophrys-Varianten van Midden-Europa” (1997).
In the Northern Italian region of Trentino, the spider orchids have lips which are are sometimes more rounded, sometimes more elongate.
At the Northern edge of the lake Garda, Monte Brione is towering, a distinctive rock massif with steep slopes on three sides. Visiting the area at the end of April you can find green meadows of spring between olive groves and the forest which is dominated by ashes, beeches and holm oaks. At a height of 170 meters, in the lower part of the mountain with a height of 376 meters, a white-flowered Orchis simia is growing, together with five violet-flowered plants of this beautiful orchid species.
In the middle of a rain shower, the monkey orchis is presenting itself in its whole beauty. The white-flowered form is quite rare. I estimate a frequency of 2 albiflora forms among 1000 plants. It’s just a genetic specialty. There is no cumulation in certain regions or biotopes which would indicate a regional evolutionary development.
In a contribution for the journal Berichte aus den Arbeitskreisen Heimische Orchideen (Peter Zschunke: Von “alba” bis “viridiflora” – zur Taxonomie und Biologie hypochromer Orchideen. In: Berichte aus den Arbeitskreisen Heimischer Orchideen 38 (1), 2021. S. 78-99) I studied the question, how albiflora forms of orchids should be addressed in a scientifically correct way. In the past, botanists have been quite inventive to describe such plants with different names and on different taxonomic levels – even up to subspecies. But there is “no need to introduce a taxon in case of an individual genetic disorder as it is the case when the development of Pigments is disturbed”. Instead I propose, “to address all individuums of orchid species with a visible genetic disorder of the development of pigments as albiflora forms, with no need of a formal description while using the most common term in descriptions and with a general understanding of the term form as a ‘nomen nudum’, which does not require a typus.”
One year after the publication of a new edition of “Orchideen in Hessen” by Heinrich Blatt, released by the Arbeitskreis Heimische Orchideen (AHO) Hessen, a first monograph about two German states in the neighborhood has been published: “Die wildwachsenden Orchideen in Rheinland-Pfalz und im Saarland“. In Coblence, six of the eight authors took part in the first presentation of the new book, honoring the 40-year-anniversary of AHO Rheinland-Pfalz/Saarland. Among them was the chairman of the AHO club, Juergen Passin (3. from right).
The book about Rhineland-Palatinate also shows several albiflora forms as Anacamptis morio, Anacamptis pyramidalis, Dactylorhiza incarnata, Dactylorhiza maculata, Dactylorhiza majalis, Gymnadenia conopsea, Orchis mascula, Orchis militaris and Orchis purpurea. The authors address those as varieties. It would be botanically more correct to name them as forms. There is also no taxonomic consistency – sometimes they are named var. albiflora, sometimes they are named var. alba. The hypochrome Ophrys forms of apifera and fuciflora are called var. flavescens, those of insectifera var. ochroleuca.
For me, it’s a bit odd that Dactylorhiza fuchsii is missing – those plants are subsumed among Dactylorhiza maculata agg. The authors are arguing that both taxa in Rhineland-Palatinate are “mixing”- while the populations in Saarland are following the “fuchsii type”. The book mostly shows photos shot in the Eifel region. There, the plants are tending to Dactylorhiza maculata with a distribution focus in Northwestern Europe. But further to the East, along the Rhine and in the Westerwald region there are also typical Dactylorhiza fuchsii.
The presentation of the book combined with a convention of AHO members was a good opportunity for professional discussions – here with Karel Kreutz (left) und Werner Hahn (right).
A witty lecture by the authors and photographers Jean Claessens und Jacques Kleinen was concluding the Day with a presentation of very detailled macro photos and the precise interaction of orchid flowers and pollinators.
The diversity of the orchid flora in Romania is presented by Nora De Angelli and Dan Anghelescu in their book The Orchids of Romania. The richly illustrated book also shows 21 taxa with albiflora forms – mainly in the genera Anacamptis, Dactylorhiza and Orchis. Four white-flowered orchids are documented as special varieties: Dactylorhiza fuchsii var. albiflora, Dactylorhiza incarnata var. albiflora, Dactylorhiza maculata var. albiflora and Epipactis palustris var. albiflora. The book shows an abundant selection von photos, selected with love and passion, among them many with visiting or pollinating insects. In Romania there are 71 orchid species and 7 further newly detected species, the authors, father and daughter, are writing. Many of them are still well represented by numerous populations, though there are different dangers threatening their existence. We have a moral obligation to protect these wonderful yet extremely vulnerable plants, Nora De Angelli and Dan Anghelescu are stressing.
Nora De Angelli also sent me some photos of albiflora orchids in Romania to be presented here, which is really great. The first to be shown are those of Dactylorhiza maculata subsp. transsilvanica which are of special interest for the understanding of albiflora orchids between spontaneous mutations and the miracles of evolution.
Himantoglossum hircinum and Himantoglossum robertianum belong to the most common orchid species in southern Europe. In case of both species there are regularly plants with flowers lacking the Anthocyanine pigment. Those albiflora forms have green and white flowers instead of flowers with reddish stripes and points.
photo: Matthias Svojtka, 4.6.2014, in Vienna
Matthias Svojtka of the University of Vienna has also found an anthocyanine free form of the much rarer Adriatic lizard orchid (Himantoglossum adriaticum). “At this Himantoglossum location in the Lower Lobau there is a quite big population of about 40 to 60 plants in open grasslands, every year there are also one or two white-flowered plants“, he wrote about his photos.
A region in the Steiermark, in the Austrian Alps, is the aim of a second hike in the mountains in Corona year 2020. On the Tauplitz-Alm I am mainly interested in the diversity of Nigritella species. Among the most common orchids of this mountain region is the fragrant orchid (Gymnadenia conopsea). Hundreds of them are flowering on meadows and forest clearings. Hiking to the Lawinenstein (1965 m) on July 7th I was surprised by a white-flowered Gymnadenia conopsea, with a long inflorescence. Gymnadenia conopsea is an orchid species which is developing white-flowered mutations quite regularly. But in most cases there are only singular plants of this form.
In most regions of its distribution area Anacamptis palustris (synonyms: Orchis palustris, Paludorchis palustris) is highly endangered, because fens and wet meadows are more and more reduced. In a fen in the Bavarian district of Rosenheim there are still growing thousands of this special species.
On a meadow with about 100 flowering Anacamptis palustris there are also 7 albiflora forms – that’s a much higher percentage than what you could expect just by random genetic mutations. H. Kretzschmar, W. Eccarius and H. Dietrich (Die Orchideengattungen Orchis, Anacamptis, Neotinea. 2007) note: Rarely there are white-flowered plants showing a photo of an albiflora form from Lake Neusiedle in Austria. It seems that white flowered Anacamptis palustris are really rare, similar to its sister species Anacamptis laxiflora.
All flowers of the albiflora plants have not the slightest hue of purple. Even the pollinia are bright with a slight yellow influence, similar to the buds.
Anacamptis palustris in this fen is accompanied by smaller groups of Orchis majalis, Dactylorhiza incarnata subsp. ochroleuca, budding Epipactis palustris and Liparis loeselii.
Among a population of about one thousand Orchis pauciflora in Southern Croatia, Bariša Ilić from Metković found 8 plants with almost white flowers. Their bright lips are contrasted with its reddish spots. This is the first time that he observed those special forms at a location near the town of Ston, in Dubrovnik-Neretva county. Bariša Ilić is an ornithologist, who fell in love with native orchids five years ago, as he explains.
Orchis pauciflora – photo: Bariša Ilić
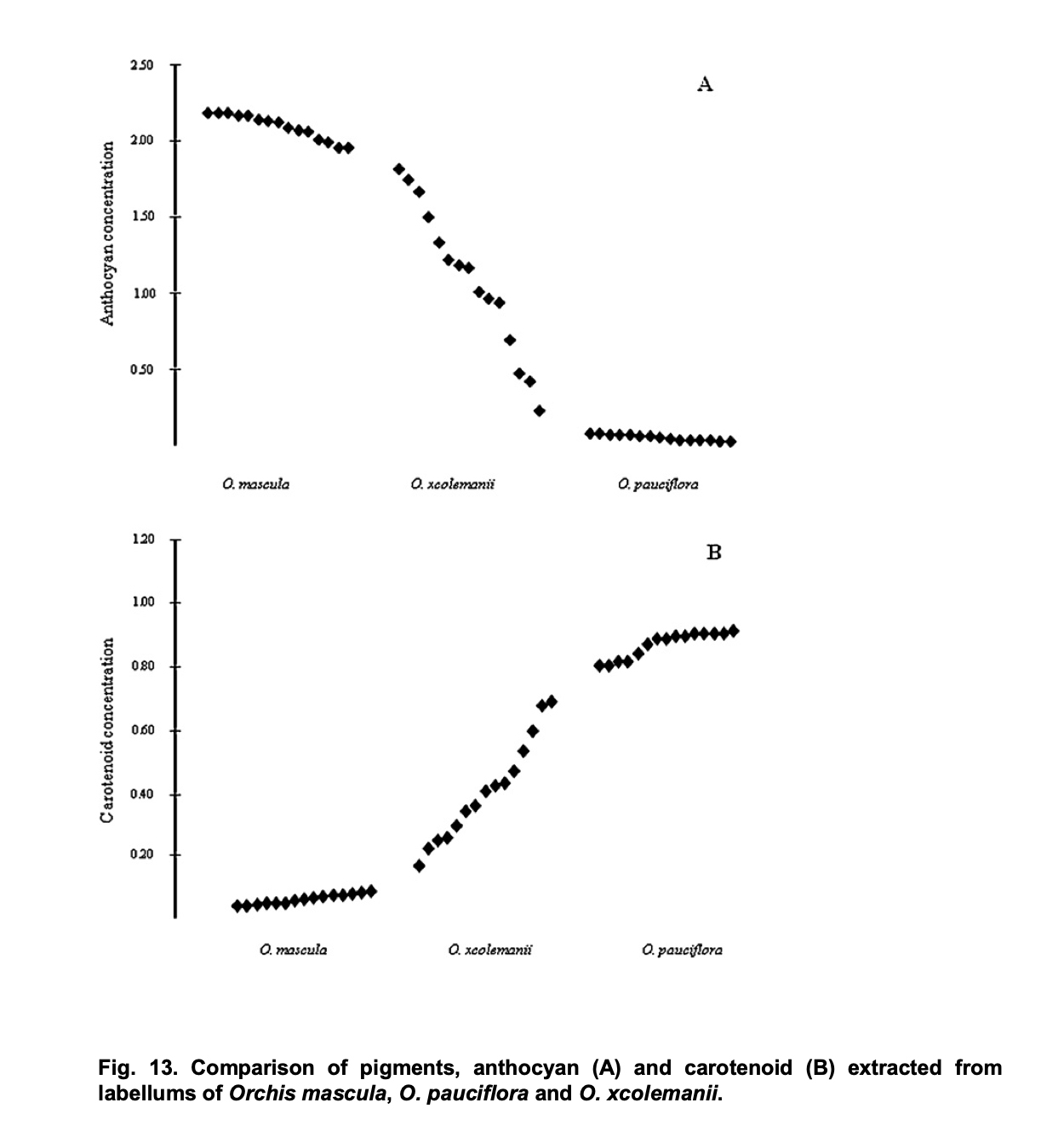
The flowers of Orchis pauciflora have a strong yellow color. They contain high concentrations of carotenoids – pigments who create the yellow flower color with many plants. While anthocyanines – which create red, purplish or brownish flower colors – are water soluble, carotenoids are lipid soluble (like chlorophyll). Since anthocyanines are produced in a complex process of biosynthesis with the participation of more than five enzymes, it is more likely that there will be a loss of anthocyanines than it might be in the case of carotenoids. This might explain why Orchis pauciflora with whitish instead of yellow flowers are extremely rare – and even in these findings, the plants seem to have retained a last rest of yellow color in the center of the labellum.
This impression of a gradual loss of carotenoid pigments corresponds with an interesting study of the Italian botanist Alessia Luca. She showed how the concentration of carotenoids is changing in the case of hybrids between Orchis pauciflora and Orchis mascula. Her thesis at the Università della Calabria includes the measuring of anthocyanin and carotenoid concentrations in the labellum of the hybrid plants, Orchis x colemanii. The results show that concentration von carotenoids is gradually decreasing in the hybrids: “O. xcolemanii showed a continuous flower color variation (Figure. 13) ranging from red-purple flowers of O. mascula to yellow flowers of O. pauciflora.” (Alessia Luca: Evolutive significance of hybridization in Mediterranean deceptive orchids, p. 42)
Alessia Luca: Evolutive significance of hybridization in Mediterranean deceptive orchids, p. 43